УДК 634.836:531.19
ВЛИЯНИЕ РЕГУЛЯТОРОВ РОСТА НА ПРОЯВЛЕНИЕ ГЕНЕТИЧЕСКИ
ДЕТЕРМИНИРОВАННЫХ ПРИЗНАКОВ ОКРАСКИ КОЖИЦЫ И СОКА ЯГОД ВИНОГРАДА НА УРОВНЕ
КАЛЛУСНОЙ ТКАНИ IN VITRO
Зленко В.А. – к. б. н.
Институт винограда и вина «Магарач» УААН
Котиков И.В. – ст. науч. сотр.
Государственный Никитский ботанический сад
Трошин Л.П. – д. б. н., профессор[1]
Кубанский государственный аграрный университет
Корреляция между синтезом антоцианов в ягодах различных сортов винограда
и в их каллусных тканях in vitro зависит от состава культуральной среды.
Генотипы винограда различаются по способности к синтезу антоцианов в каллусных
тканях, культивируемых при освещении интенсивностью 2000 лк на модифицированной
твердой среде Schenk, Hildebrandt (1972) в зависимости от концентрации
в ней регуляторов роста НУК (NAA) и БАП (BAP). В варианте среды с высоким
содержанием БАП (8 мг/л) и 2 мг/л НУК у сорта Антей магарачский образовался
зеленый каллус, а у Муската черного – красный. Наоборот, добавление в
эту среду высокой концентрации НУК (10 мг/л) и 0,5 мг/л БАП индуцировало
развитие красного каллуса у Антея магарачского и серого – у Муската черного.
Вариант этой среды с 2 мг/л НУК и 0,5 мг/л БАП вызывал формирование красного
каллуса у обоих сортов. Повышение рН этого варианта среды до 7,0 (до автоклавирования)
увеличивало синтез антоцианов в каллусах названных сортов. У сеянца Магарач
100-74-1-7 максимальное накопление антоцианов в каллусе наблюдалось в
варианте этой среды с 2 мг/л НУК и низкой концентрацией БАП (0,05 мг/л).
Выведение сортов винограда занимает 10–25 лет, на протяжении которых
создают обширный гибридный фонд и в процессе его изучения выделяют по
сочетаемости необходимых градаций признаков и свойств единичные сеянцы.
Для ускорения селекционного процесса необходимо совершенствовать методы
его проведения. Одним из подходов к решению этой проблемы может стать
ранняя диагностика хозяйственно-ценностных признаков на ювенильной стадии
задолго до вступления сеянцев в пору плодоношения.
Все клетки растительного организма обладают потенциальной способностью
к биосинтезу фенольных соединений, образующихся из продуктов первичного
метаболизма [1]. Вследствие тотипотентности растительных клеток, изменение
внешних условий может вызвать вторичную дифференциацию, в частности, превращение
меристематических клеток в паренхиматические [2]. Последние бедны цитоплазмой,
имеют мелкие ядра, большие вакуоли и пластиды, наполненные запасными веществами.
Характер содержимого паренхиматических клеток тесно связан с выполняемыми
ими функциями – синтезом и накоплением различных запасных веществ. Преобладание
в каллусной ткани клеток паренхимного типа способствует синтезу и формированию
вторичных веществ [3]. Антоцианопласты содержались в вакуолях клеток кожицы
ягод, а также ее культуры in vitro [4].
Суспензия клеток винограда использовалась для определения предшественников
синтеза антоцианов. Установлено, что фенилаланин, меченный изотопом 13С,
включается в синтез антоцианов на уровне 65 % [5]. Накоплению антоцианов
в клетках суспензионной культуры Vitis spp. предшествовало повышение в
них уровня фенилаланина, из которого синтезируются антоцианы [6]. Добавление
в среду фенилаланина увеличивало синтез антоцианов [7].
Внесение в среду 0,1–10 мг/л 2,4-дихлорфеноксиуксусной кислоты (2,4-Д)
вызывало дедифференциацию клеток Haplopappus gracilis, уменьшение их размеров
с одновременным прекращением синтеза антоцианов [8]. По модельной системе
установлено, что синтез антоцианов в суспензионной культуре моркови индуцируется
переносом клеток из среды, содержащей 2,4-Д, в среду без нее [9].
В роли индукторов синтеза в клетках вторичных веществ могут выступать
различные контролируемые факторы: добавление в среду ауксинов, гиббереллинов,
цитокининов и др., температура культивирования, условия аэрации среды
и т. д. [3]. Цитокинины и их предшественники в определенных концентрациях
способствуют синтезу антоцианов [2]. Каллусы из усиков винограда сорта
Кардинал, выращенные на питательных средах, содержащих кинетин, гиббереллин
и аденин в определенных соотношениях, приобретают пигментацию от красного
до фиолетового цвета [10].
Обработка абсцизовой кислотой гроздей винограда в начале созревания (10
% проявления окраски ягод) на 3–4 недели ускоряла процесс созревания ягод
и синтеза в них антоцианов [11]. При воздействии эндогенной абсцизовой
кислотой на ягоды винограда в их эпидермальных клетках увеличивалось образование
и размер антоцианопластов без изменения размера клеток [12].
В культуре каллусной ткани Parthenocissus tricuspidata Planch. при уменьшении
концентрации азота синтезировались антоцианы [13]. Снижение уровня накопления
иона NO3- и увеличение содержания сахарозы в среде
ускоряли синтез антоцианов в клетках суспензионной культуры винограда.
Максимальный синтез антоцианов отмечался при 2,5 mM NO3-
и 8 % сахарозы. Ионы NO3- ингибируют аккумулирование
антоцианов с помощью тонопласт АТФ-азы [14]. В суспензионной культуре
сорта винограда Gamay Freaux синтез антоцианов наблюдался на пятый день
культивирования и был увеличен соответственно на 32 и 46 % в среде среднего
(0,25 mM) и низкого (0,008 mM) уровней содержания неорганического фосфата
[15].
В отличие от сорбитола и манитола, высокие концентрации сахарозы, глюкозы
и фруктозы в суспензионной культуре винограда стимулировали синтез антоцианов.
По мнению авторов, увеличение синтеза антоцианов связано не с повышением
осмoтического потенциала в культуральной среде, а с включением сахаров
(сахарозы, глюкозы и фруктозы) в метаболические процессы [16].
Если повторные деления клеток тормозятся, и нарастание клеточной массы
прекращается за счет факторов, лимитирующих условия культивирования, в
клетках наблюдается повышенный синтез антоцианов [17]. В суспензионной
культуре Vitis spp. антоцианы аккумулируются на стационарной фазе, когда
клеточные деления уже прекращаются. Ингибирование синтеза ДНК афидиколином
или добавление свежей жидкой среды без фосфата тормозит процесс клеточного
деления и приводит к синтезу антоцианов [7].
Синтез антоцианов зависит от состава света и температуры. Освещение с
преобладанием зеленых и синих участков спектра способствовало появлению
антоцианов в каллусной ткани винограда. Обработка кусочков каллусной ткани
корня моркови холодной водой (+ 4оC) перед высадкой ее на питательную
среду также приводила к синтезу антоцианов [2].
На основании литературных данных можно сделать вывод о том, что синтез
антоцианов в каллусной ткани зависит от состава питательной среды, освещения
и температуры. Образование и накопление антоцианов увеличивается в стационарной
фазе роста.
На среде, способствующей синтезу антоцианов, выделенные клоновой селекцией
две клеточные линии сорта Gamay синтезировали в 4 раза больше антоцианов,
чем исходные клетки. Качественный состав антоцианов менялся в процессе
культивирования и зависел от условий выращивания культуры [18].
В каллусной ткани петунии содержались те же антоцианы (петунидин и мальвидин),
что и в венчиках исходных растений [19]. Состав культуральной среды влияет
на количественный состав антоцианов и их предшественников в клетках винограда
in vitro. Относительно контрольных клеток, на средах с невысокой концентрацией
неорганического фосфата аккумулировалось больше ацилированных цианидин-
и пeонидин-глюкозидов [15]. Пролиферирующие каллусные клетки винограда
сорта Spätburgunder, культивируемые in vitro, синтезировали продельфинидин
[20].
Вещества антоцианового комплекса, углеводы и ароматические вещества накапливаются
в ягодах винограда по мере их созревания. Максимум формирования этих веществ
наблюдается на стадии физиологической зрелости винограда, когда мякоть
ягод состоит из паренхиматических клеток с большими вакуолями, где содержатся
углеводы, антоцианы и ароматические вещества [21]. В культуре ткани при
определенных условиях можно вызывать образование каллусной ткани паренхиматического
типа [2]. Нами была выдвинута гипотеза о возможности получения каллусной
ткани in vitro паренхиматического типа, приближающейся по своему строению
к мякоти ягоды на стадии физиологической зрелости.
Разработан способ ранней диагностики окраски кожицы и сока ягод у сеянцев
(в первые месяцы их развития) на уровне каллусной ткани, выращенной in
vitro. У сортов с белой кожицей и соком ягод, образующимся из эксплантов
листьев, черешков листьев и междоузлий, формируется каллус белого цвета.
У сортов с окрашенной кожицей, но с бесцветным соком красный каллус будет
только при его культивировании с интенсивностью освещения 2000 лк, а у
сортов с окрашенными кожицей и соком ягоды – как в темноте, так и при
освещении [22, 23].
У винограда генотипическая специфичность сорта или сеянца по содержанию
и порядку синтеза веществ – компонентов антоцианового комплекса (диглюкозидов,
в частности) может быть установлена на уровне каллусной ткани. Выявленная
корреляция между содержанием и порядком синтеза антоцианов в кожице ягод
и каллусе, развившемся из эксплантатов (междоузлий, листьев, черешков
листьев), позволяет селекционерам проводить раннюю диагностику этих признаков
в будущем урожае сеянцев винограда в первые месяцы их развития [24].
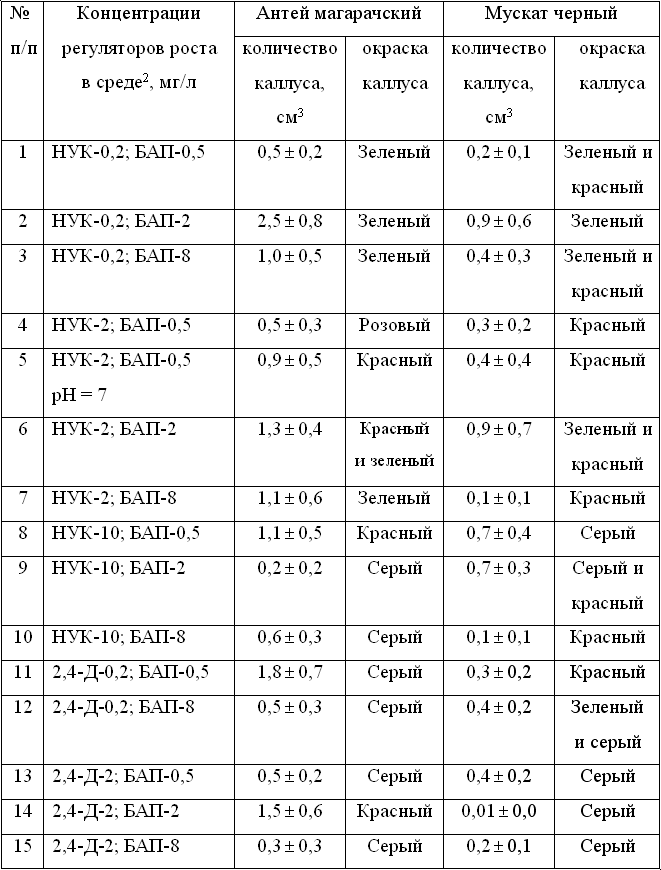
Генотипы винограда различаются по ответной реакции каллусной ткани, культивируемой
in vitro, на концентрации регуляторов роста в питательной среде (табл.
1).
Таблица 1. Различия двух сортов винограда по развитию
и окраске каллусных тканей in vitro на вариантах сред с различными концентрациями
регуляторов роста НУК, 2,4-Д и БАП 1
1 Первичные экспланты – черешки листьев, 30 дней культивирования
в темноте и затем 28 дней при освещении интенсивностью 2000 лк белыми
люминесцентными лампами, 15 повторностей, Р < 0,05.
2 Среда [25] с повышенной концентрацией сахарозы (60 г/л),
7,5 г/л Difco агара, рН = 5,6 (кроме варианта среды № 5).
Низкая концентрация (0,2 мг/л) ауксина 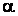 -нафтилуксусной
кислоты (НУК) с 0,5 и 2 мг/л 6-бензиламинопурина (БАП) не способствовала
синтезу красящих веществ в каллусах винограда сортов Антей магарачский
и Мускат черный. Добавление в среду высокой концентрации НУК (10 мг/л)
с 0,5 мг/л БАП вызывало развитие красного каллуса Антея магарачского и
серого – Муската черного, наоборот, содержание в среде 8 мг/л БАП с добавлением
2 мг/л НУК приводило к образованию красного каллуса у Муската черного
и зеленого – Антея магарачского. На среде, содержащей другой ауксин –
2,4-дихлорфеноксиуксусную кислоту (2,4-Д) низкой концентрации (0,2 мг/л)
с 0,5 мг/л БАП, развивались красный каллус у сорта Мускат черный и серый
– Антея магарачского. Только один вариант концентрации регуляторов роста
– 2 мг/л НУК и 0,5 мг/л БАП – индуцировал синтез антоцианов в каллусах
двух сортов винограда без их окрашивания в зеленый цвет. Высокое значение
рН = 7 среды улучшало образование красящих веществ в каллусе этих сортов
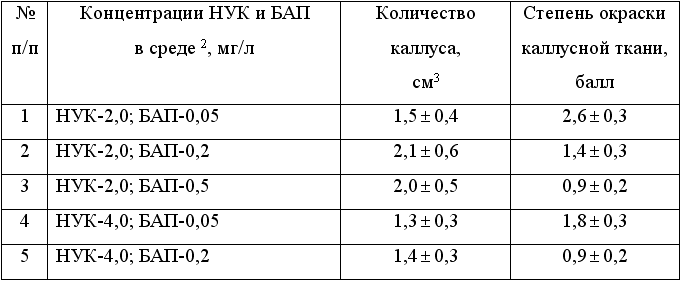
(см. табл. 1). Для синтеза антоцианов в каллусной ткани гибридной формы
Магарач 100-74-1-5 оптимальным был вариант среды, содержащий 2 мг/л НУК
и 0,05 мг/л БАП (табл. 2). Повышение концентрации этих регуляторов роста
в вариантах среды снижало синтез красящих веществ в каллусных тканях. -нафтилуксусной
кислоты (НУК) с 0,5 и 2 мг/л 6-бензиламинопурина (БАП) не способствовала
синтезу красящих веществ в каллусах винограда сортов Антей магарачский
и Мускат черный. Добавление в среду высокой концентрации НУК (10 мг/л)
с 0,5 мг/л БАП вызывало развитие красного каллуса Антея магарачского и
серого – Муската черного, наоборот, содержание в среде 8 мг/л БАП с добавлением
2 мг/л НУК приводило к образованию красного каллуса у Муската черного
и зеленого – Антея магарачского. На среде, содержащей другой ауксин –
2,4-дихлорфеноксиуксусную кислоту (2,4-Д) низкой концентрации (0,2 мг/л)
с 0,5 мг/л БАП, развивались красный каллус у сорта Мускат черный и серый
– Антея магарачского. Только один вариант концентрации регуляторов роста
– 2 мг/л НУК и 0,5 мг/л БАП – индуцировал синтез антоцианов в каллусах
двух сортов винограда без их окрашивания в зеленый цвет. Высокое значение
рН = 7 среды улучшало образование красящих веществ в каллусе этих сортов
(см. табл. 1). Для синтеза антоцианов в каллусной ткани гибридной формы
Магарач 100-74-1-5 оптимальным был вариант среды, содержащий 2 мг/л НУК
и 0,05 мг/л БАП (табл. 2). Повышение концентрации этих регуляторов роста
в вариантах среды снижало синтез красящих веществ в каллусных тканях.
Таблица 2. Влияние регуляторов роста -нафтилуксусной
кислоты (НУК) и 6-бензиламинопурина (БАП) на развитие и окраску каллусной
ткани in vitro, образовавшейся 1 из эксплантов черешков листьев
гибридной формы винограда Магарач 100-74-1-5
1 30 дней культивирования в темноте и затем 28 дней при освещении
интенсивностью 2000 лк белыми люминесцентными лампами, 15 повторностей,
Р < 0,05.
2 Среда [25] с повышенной концентрацией сахарозы (60 г/л),
7,5 г/л Difco агара, рН = 5,6.
Что касается возможности диагностирования содержания ароматических веществ
в ягодах винограда на уровне каллусной ткани in vitro, то в клетках суспензии
из тканей черешков листьев сорта Конкорд ароматические вещества, характерные
для сортов Vitis labrusca L., не найдены, а предшественники  -дамасценона присутствовали в меньшей
концентрации, чем в каллусе на твердой среде, причем только на стационарной
фазе роста суспензии. Интенсивное освещение и увеличенная концентрация
сахарозы в культуральной среде повышали уровень содержания предшественников
-дамасценона
в каллусной ткани, полученной из черешков листьев сорта Кoнкорд [26 ]. -дамасценона присутствовали в меньшей
концентрации, чем в каллусе на твердой среде, причем только на стационарной
фазе роста суспензии. Интенсивное освещение и увеличенная концентрация
сахарозы в культуральной среде повышали уровень содержания предшественников
-дамасценона
в каллусной ткани, полученной из черешков листьев сорта Кoнкорд [26 ].
Таким образом, синтез красящих веществ в каллусной ткани на специальной
среде при определенных условиях (без освещения или с освещением) позволяет
проводить раннюю диагностику содержания антoцианов в кожице и соке ягод
сеянцев в первый год их развития, задолго до плодоношения [22], при этом
можно прогнозировать качественный состав веществ антоцианового комплекса
[24].
Список литературы
1. Запрометов М.Н. Фенольные соединения растений: биосинтез, превращения
и функции // Новые направления в физиологии растений. – М.: Наука, 1985.
– C. 143–162.
2. Бутенко Р.Г. Культура изолированных тканей и физиология морфогенеза
растений. – М.: Наука, 1964. – 270 с.
3. Калинин Ф.Л. Методы культуры тканей в физиологии и биохимии растений
/ Ф.Л. Калинин, В.В. Сарнацкая, В.Е. Полищук. – Киев: Наукова думка, 1980.
– 488 с.
4. Nakamura M. Anthocyanoplasts in Kyohko grapes // Journal of the Japanese
Society for Horticultural Science. – 1993. – V. 62. – P. 353–358.
5. Production of 13C–labelled anthocyanins by Vitis vinifera
cell suspensions cultures / S. Krisa, P. Waffo Teguo, A. Decendit, G.
Deffieux, J. Vercauteren, J.M. Merillon // Phytochemistry. – 1999. – V.
51. – P. 651–656.
6. Obtaining Vitis vinifera cell cultures producing higher amounts of
malvidin-3-0-β-glucoside / S. Krisa, X. Vitrac, A. Decendit, F. Larronde,
G. Deffieux, J.M. Merillon // Biotechnology Letters. – 1999. – V. 22.
– P. 497–500.
7. Regulatory mechanisms of biosynthesis of betacyanin and anthocyanin
in relation to cell division activity in suspension cultures / M. Sakuta,
H. Hirano, K. Kakegawa, J. Suda, M. Hirose, R.W.IV Joy, M. Sugiyama, A.
Komamine // Plant Cell, Tissue and Organ Culture. – 1994. – V. 38. – P.
167–169.
8. Stiсkland K., Sunderland N. Production of anthocyanins, Flavonols
and Chlorogenic Acids by Cultured Callus Tissues of Haplopappus gracilis
// Annals of Botany. – 1972. – V. 36. – № 146. – P. 443–457.
9. Ozeki Y., Komamine A. Induktion of anthocyanin synthesis in relation
to embryogenesis in a carrot suspension culture – a model system for the
study of expression and repression of secondary metabolism // Primary
and secondary Metab. Plant Cell Cult. – Berlin e. a., 1985. – P. 99–106.
10. Изровска Н., Лилов Д. Участие на гиберелиновата киселина / ГАЗ/
при индуциране на морфогенез върху тъкани, изолирани от различни органи
на лоза // Физиология на растенята. – 1981. – T. 4. – № 1. – С. 23–28.
11. Effects of natural type (S)–(+)–abscisic acid on anthocyanin accumulation
and maturity in ‘Kioho’ grapes / K.S. Lee, J.C. Lee, Y.S. Hwang, I.B.
Hur // Journal of the Korean Society for Horticultural Science. – 1997.
– V. 38. – P. 717–721.
12. Kim S.B. Effect of ABA on the occurrence and development of anthocyanoplasts
in ‘Kyoho’ grapes / S.B. Kim, C.H. Lee, D.H. Han // Journal of the Korean
Society for Horticultural Science. – 1997. – V. 38. – P. 55–59.
13. Heller R. Sur la formation d’anthocyane dans les tissus de vigne
– vierge cultive in vitro par supperssion de l’azote du milieu de culture
// C. R. Soc. Biol. – 1948. – V. 142. – P. 768–769.
14. Enhanced anthocyanin production in grape cell cultures / F.J. Hirasuna,
M.L. Shuler, V.K. Lackney, R.M. Spanswick // Plant Sciense (Shannon).
– 1991. – V. 78. – P. 107–120.
15. Dedaldechamp F. Enhancement of anthocyanin synthesis and dihidroflavonol
reductase, DFK, activity in response to phosphate deprivation in grape
cell suspensions / F. Dedaldechamp, C. Uhel, J.J. Macheix // Phytochemistry.
– 1995. – V. 40. – P. 1357–1360.
16. Regulation of polyphenol production in Vitis vinifera cell suspension
cultures by sugars / F. Laronde, S. Krisa, A. Decendit, C. Cheze, G. Deffieux,
J.M. Merillon // Plant Cell Reports. – 1998. – V. 17. – № 12. – P. 946–950.
17. Enhanced accumulation of anthocyanins in Vitis vinifera cells immobilized
in polyurethane foam / J.L. Iborra, J. Guardiola, S. Montaner, M. Canovas,
A. Manjon // Enzyme and Microbial Technology. – 1994. – V. 16 – P. 416–419.
18. Cormier F. Anthocyanin production in selected cell lines of grape
Vitis vinifera L. / F. Cormier, C.B. Do, Y. Nikolas // In vitro Cellular
and Developmental Biology – Plant. – 1994. – V. 30. – P. 171–173.
19. Synthesis of malvidin and petunidin in pigmented tissue cultures
of Petunia hybrida / C.M. Colijn, L.M.V. Jonsson, A.W. Schram, A.J. Kool
// Protoplasma. – 1981. – V. 107. – № 1-2. – P. 63–68.
20. Feucht W. Flavanols in grapevine: in vitro accumulation and defence
reactions in shoots / W. Feucht, D. Freutter, E. Christ // Vitis. – 1996.
– V. 35 – P. 113–118.
21. Стоев К.Д. Физиологические основы виноградарства. – София: Изд. Болгарской
акад. наук, 1973. – Т. 2. – 538 с.
22. А. с. 948339, СССР, МКИ3 АО1G 7/00. Способ диагностики
окраски кожицы и цвета сока ягод винограда. – Заявка № 3232084/30-15.
Заявл. 04.01.81. Опубл. 07.08.82. / П.Я. Голодрига, В.А. Зленко, Т.А.
Калуцкая, Л.К. Киреева, Н.П. Дубовенко, Р.Г. Бутенко, Б.А. Левенко. –
Бюл. № 29. – 2 с.
23. Haydu Z. Application of in vitro cultures in the Research on grapevine
genetics // 4-th international symposium on grapevine breeding. – Verone
/Italie/ 13–18 Avril, 1985. – P. 25.
24. Голодрига П.Я. Прогнозирование некоторых компонентов качества урожая
сортов и сеянцев винограда по каллусной ткани / П.Я. Голодрига, М.А. Костик,
В.А. Зленко // Физиология и биохимия культурных растений. – 1986. - №
5. – С. 510–515.
25. Schenk R.U., Hildebrandt A.C. Medium and techniques for induction
and growth of monocotyledonous and dycotyledonous plant cell cultures
// Can. J. Bot. – 1972. – V. 50. – P. 199–204.
26. Shure K.B., Acree T.E. Production of β–damascenone precursors
in cell cultures of Vitis labruscana cv. Concord grapes // Plant Cell
Reports. – 1994. – V. 13. – P. 477–480.
| 
 English
English


 Версия для печати
Версия для печати Файл в формате pdf
Файл в формате pdf